地球最早的生命除了细菌之外,还有古细菌。
古细菌
古细菌(又可叫做古生菌、古菌、古核细胞或原细菌)与真细菌几乎同时出现于40亿或者38亿年前的生命诞生之初(实质是同时被造)。
它也是单细胞的无核生物,有球形、杆形、螺旋形、叶状、方形等各种形状。
单个古菌细胞直径在0.1到15微米之间,有一些种类形成细胞团簇或者纤维,长度可达200微米。
很多古菌生存在极端环境中:
一些生存在沸腾的热泉或者海底黑烟囱中;
一些生存在很冷的环境如极地冰盖下;
许多生存在高盐、强酸或强碱性的水中;
也有些古菌是嗜中性的,能够在沼泽、废水和土壤中生活;
很多产甲烷的古菌,生活在动物的消化道中。
1983年,科学家们在太平洋中抽取洋底的水样时发现,从地球内部迸发出来的热气和岩浆还在沸腾,一群古细菌却正在那里洗热水澡,不禁大吃一惊。
他们是了不起的生命,挑战极限,蔑视教条,十分顽强。
有些极端嗜热菌只能生长在90℃以上的高温环境,80℃以下即失活;
意大利海底的一族古细菌,降至84℃即停止生长;
一些火山口附近的古菌甚至可以生活在250℃左右的环境中;
极端嗜盐菌耐受盐度可达25%(如死海和盐湖);
极端嗜酸菌能生活在pH值1以下的地方;
极端嗜碱菌多数寄居在碱湖、碱池中,pH值可达11.5以上;
盐杆菌可以利用光能制造ATP(尽管不像其他利用光能的生物一样利用电子链传导实现光合作用);
……
图/生存在海底热泉喷口的Geogemma barossii细菌 来源/Medical University of Graz, Austria
死海中生活着许多耐盐古细菌:
图/嗜盐菌(来源 中国科普博览)
据科学家推测,按照换算成碳元素来衡量,整个地球海底之下地层中的古细菌总量可达900亿吨,相当于陆地土壤中各种微生物总量的3倍以上。
古细菌有扁平直角几何形的细胞,有独特的辅酶,许多古细菌有内含子,膜含二醚而不是酯(膜脂由甘油醚构成,是为了适应超高温环境),没有基因调节机制,喜高温酷寒,许多严格厌氧,都与真细菌有所不同。其鞭毛的成分和形成过程也与真细菌路径不一,在复制、转录和翻译上更是接近真核生物,弄得进化派的生物学家们如坠五里雾中。
受达尔文误导,大多数科学家们相信,在漫长的进化过程中,每种生物细胞中的信息分子(核酸和蛋白质)的序列均不断发生着突变,许多信息分子序列的变化在时间上是随机的,进化速率相对恒定。
这些基因或其产物乃是记录生物进化历程的分子记时器——物种间的亲缘关系,可以用它们共有的某个具有时钟特性的基因或其产物(如蛋白质)在序列上的差别来加以定量描述。
习惯从进化角度去看问题,使得几种最早生物之间的关系变得错综复杂,惝恍迷离。
把古细菌从原核生物中划出来另立一个古细菌界,与原核生物、真核生物并列,实际上并不科学。
基于rRNA序列的所谓系统发生树,认为细菌、古菌和真核生物各自代表了一支具有简单遗传机制的远祖生物的后代、三者同祖,更是无稽之谈。——伍斯等人以古菌命名之,反映了这一幻想(希腊语archae为“古代的”)。
说穿了,古菌其实就是高级生命制造出来,以适应在冥古宙、太古宙极端环境生活的一种生物。
那时的地球大气中充满了有毒气体,为了生活于高温、高盐和缺氧之类的环境,古菌的生理机制、化学组成当然要很特殊。
古菌根本也谈不上什么进化,四十亿年基本上一仍旧贯(当然,生物都有弹性,古菌也不例外)。
正如网友所说:
32万8千亿代“进化”下来,细菌还是细菌,没有变异成别的物种。
说明了基因的超稳定性,已经在事实上否定了“进化论”。
古菌研究正在世界范围内升温,科学界认为古菌中蕴藏着远多于另两类生物的、未知的生物学过程和功能,有助于阐明生物进化规律的线索。
然而微生物基因组测序令科学家们极度迷惑,不知所之。他们主观假想出来的进化树,总是难以枝杈分明、线路清晰。
自诞生之日起,伍斯的“三域学说”便遭到部分人,特别是微生物学领域外学者的反对。
反对者坚持认为:
原核与真核的区分,是生物界最根本的、具有进化意义的分类法则(呵呵!);
与具有丰富多样性表型的真核生物相比,古菌与细菌的差异远没有大到需要改变二分法则的程度。
譬如詹氏甲烷球菌的蛋白质,就是44%像细菌,只有13%接近真核生物。
在另一个古菌,“嗜热碱甲烷杆菌”的基因组中也有类似情况。
从基因组比较的数字上看,古菌与细菌间的差异远小于古菌与真核生物间的差异。
概而言之,伍斯等人的论据还不足以说服“三域学说”的反对者们,“三域学说”在生物历史上恐怕将会是昙花一现。
最初的原始生命进行无氧呼吸,大多通过分解攫取的氨基酸、糖类及脂肪获取能量;有些依靠无机氧化物的还原获取能量(化能合成);有些通过光合作用摄取太阳能,如紫硫细菌、绿硫细菌、太阳杆菌等。它们以硫化氢、硫或氢气等等作为电子供体,不产氧,只拥有光合系统I(蓝藻则拥有光合系统I和光合系统II)。
细菌之类原核细胞已具有相当复杂的结构和功能。
用电子显微镜观察,已报道的细菌超微结构有细胞壁、胞浆膜、鞭毛、纤毛、荚膜、核糖核蛋白颗粒、中膜小体、核区、芽胞等。进一步还观察到了一些真核细胞才有的细胞器,如微管、微丝、板层样结构等。膜中存在着大量不同的蛋白质,具有非常多的生理功能,集中执行着多数代谢功能:
如构成了细胞的渗透屏障;
主动运输,控制物质的流入和流出;
附着细胞色素、铁硫蛋白、ATP合成酶和其他组分;
进行细胞壁和荚膜、磷脂的生物合成以及分泌胞外酶;
附着光合作用膜、固氮酶;
提供鞭毛的着生位点,提供细菌运动能量;可能还附着细菌DNA的复制中心……等等。
蓝藻
紧接着细菌和古细菌出现的蓝细菌(蓝藻),据说已经有36亿年的历史,也有距今35 - 33亿年的说法(根据非洲、加拿大、澳大利亚等处化石)。
蓝藻,又称蓝细菌、蓝绿菌或蓝绿藻。
传统上归于藻类,后来发现它与细菌非常接近,已被归入细菌域,但国内的许多教科书上还没有改(所有的蓝藻都含有一种特殊的蓝色色素,“蓝藻”因此得名)。
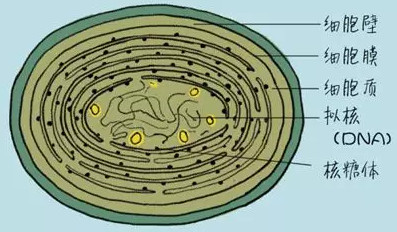
图/蓝藻细胞结构
蓝藻是单细胞原核生物,细胞壁分内外两层。唯一的有形细胞器是核糖体。在蓝藻中还有一种环状DNA——质粒,在基因工程中担当了运载体的作用。
蓝藻含有叶绿素和类胡萝卜素、藻蓝素,部分种类含藻红素。
红海就是由于水中这种蓝藻甚多,使海水呈现出红色。它的身体不具鞭毛,藻体有单细胞体的、群体的和丝状体的。
蓝藻虽无叶绿体,但在电镜下可见细胞质中有很多光合片层,叫类囊体,各种光合色素均附于其上,光合作用过程在此进行。贮藏的光合产物主要为蓝藻淀粉和蓝藻颗粒体等。它的细胞膜上还有进行有氧呼吸的相关色素和酶。
蓝藻拥有光合系统I和光合系统II(与绿色植物、藻类一样),光合作用机制较化能合成细菌要完善、复杂的多(但不是进化而来)。这在生物装备上是一个空前的大跃进。
蓝藻是最早的光合放氧生物(自养生物能够利用太阳光能,以无机物为原料合成有机物,并放出氧),被认为对地球表面从无氧大气变为有氧大气起了巨大作用(外国有科学家认为火山的作用更大)。
有不少蓝藻(如鱼腥藻),可以直接固氮(含有固氮酶),提高土壤肥力。有的蓝藻为食品,如著名的发菜和普通念珠藻(地木耳)、螺旋藻等等。
蓝藻等藻类,也是鲢鱼、鳙鱼的食物。
常见的蓝藻,有蓝球藻(色球藻)、念珠藻、颤藻、发菜等。
主要分布在有机质较多的淡水中;
部分生于海水中,潮湿、干旱的土壤或岩石上、树干和树叶上,温泉中、冰雪上;
甚至盐卤池、岩石缝中都可以发现蓝细菌的踪迹;
有些还可穿入钙质岩石或介壳、土壤深层;
也有同真菌共生形成地衣,或生于植物体内形成内生植物的。
蓝藻已知约2000种,中国已有记录的约900种,分布十分广泛。淡水水域污染,富营养化会引起蓝藻大爆发,会产生有腥臭味的“水华”,进一步污染饮用水源,亦会令鱼类缺氧死亡。

图/昆明池蓝藻爆发
蓝藻既能进行光合作用,又可以进行有氧呼吸,非常值得注意(今天的大多数原核生物既能进行有氧呼吸也能进行无氧呼吸,有氧呼吸的效率是无氧呼吸的18倍)。
“光合作用”和“有氧呼吸”的机制究竟有多么复杂,我们很快就会看到。

图/太湖蓝藻
“有氧呼吸”与“光合作用”哪一个发生在先?
科学家们说法不一。
许多人认为,光合作用应该发生在原始生物中分化出蓝藻,蓝藻制造出足够的氧气之后。没有氧气,怎么光合?
大多数人则认为,光合作用的历史更加久远。
最初的某些细菌已经在通过光合作用摄取太阳能(有些含有卟啉类化合物),只是它们装备简陋,仅仅拥有光合系统I罢了。
蓝藻出现之前地球上也不是绝对无氧,强烈的紫外线对水的光解,富含氧气分子的岩石因火山活动释放氧气…等等,都会制造出数量可观的自由氧。只是氧气这种东西过于活泼,水性杨花,她们很快就与其它物质发生了结合。
用于呼吸作用的细胞色素C排列在细胞的外表面上,而ATP合成酶则位于内膜上。所有生物细胞质膜的这种固定的方向性也被认为必定是起源于外在的有氧环境,绝大多数原核生物都能进行有氧呼吸似乎也加强了这种说法的说服力。
服膺创造论,解读就十分简单:
上天为了种种目的,把蓝菌投入地球而已。
它一开始就拥有复杂的呼吸、光合系统,对自由氧的劫持能力较之别人也毫不逊色。