在《达尔文的黑匣子》一书,贝希教授不仅讨论了细菌鞭毛,还列举了细菌纤毛、细胞内运输、免疫系统、合成核苷酸等几十个例子,让许多进化论者穷于应对。
譬如说:
一、血液凝固系统
血液凝固系统是一个串级系统:一个部件激活另一个部件,这个部件再激活第三个部件,依次类推,这样一个连环扣系统毫无疑问具有不可约化的复杂性。
在这种环环相扣中,任何一个环节出了问题,整个系统就坍塌了。
血液凝固系统就是这样一个由纤维蛋白原、凝血酶、凝血素等等蛋白质组成的自我催化琏。而激活司徒因子(一种凝血蛋白)的又是两条不同的串级途径。除此之外,整个系统还需要一个停止机制,以免全身的血液都被凝固。这个完成血凝块的形成、限制、加强和去除的是一整套完整的生物系统,各部件之间配合到如此的丝丝入扣,以至于任何一个环节的问题都会给人体造成极有可能致命的疾病。——这些零件能单独形成吗?
按照基因突变速率,据估计,其中一个蛋白(TPA)产生的几率是1/10的18次幂,至少需要100亿年才能发生。如果同时进化出和它相互作用的蛋白,几率就是1/10的36次幂。
“血液凝固系统”由进化而来的可能,在概率上根本就不存在。
二、细胞内运输系统
生物细胞内大部分的蛋白质都是在细胞核内生产出来,再被分送到各自需要的地方去。这就需要一个象邮政投递系统那样的运输系统,这必须包括这些功能:
货物被标上正确的地址;
运输器正确地识别地址并把货物装载上正确的交通车;
当交通车到了目的地时它必须知道;
以及货物被下载。
单就运输部分而言,细胞内参与其中的就包括细胞核、DNA、RNA、细胞质、SRP(信号识别颗粒)等等若干复杂的亚细胞结构。
而下载的方式通常还有三种:
门输运、
类似的膜间输运、
以及更复杂一些的囊输运。
门输运和类似的膜间输运至少需要三个部件:标志号、扫描器、以及由扫描器控制的门。
而囊输运则需要六个部件:车上的标志号、装载车的卡车、卡车上的扫描器、卡车上的标志号、仓库的扫描器、以及可控仓库大门。
用生物语言,这对应于mannose-6-phosphate、the clathrin vesicle(甘露糖-6-磷酸网格蛋白泡)中的M6P接受器、v-SNARE、t-SNARE、(v-SNARE——突触囊泡蛋白,t-SNARE——突触前膜上的靶蛋白)以及SNAP/NS蛋白质(SNAP是可溶性NSF附着蛋白。NSF是N-乙基马来酰亚胺敏感因子)等。
仔细分析这样一个环环相扣的系统,也难以使人相信这是由一些新、旧(曲线进化)部件偶然碰在一起产生的。
三、免疫系统
免疫系统是如此的重要,且功能完美,以至吸引了全世界上千的实验室进行研究。
贝希在书中描述了三个方面:无性选择、抗体多样性、以及辅助系统。
即使在简化的无性选择机制中,至少也需要这样三种部件:膜固定形态抗体、信使、以及输出形态抗体。
而且使得抗体上亿种的多样性成为可能的DNA断裂和重组机制也至少需要三类部件:基因本身、基因段起始和终了的信号、以及识别这种信号的分子生物机制。
最后,对抗体标志好的入侵者完成捕杀工作的辅助系统又是一个串级系统,所有这些都显然具有不可约化的复杂性。
细胞生物学层次上的比较免疫学帮不上什么忙,因为除了没有深入到分子生物的层次以外,这种比较学的研究本质上只提供分类,而不是如何起源的证据。
四、核苷酸合成系统
前面三个例子是明显具有不可约化复杂性的系统,渐进进化的困难也是非常明显的。
贝希下面给了一个,乍一看似乎可以用“渐进进化”解释的例子。然而,仔细考察合成的细节就会发现,问题远不是那么简单。
这就是“合成核苷酸”的反应:
DNA有四种核苷酸基本结构单位,分别称作A 、C 、G 、T 。A 在和聚合物联结以前,可以有几种形态,包括AMP、ADP、ATP。单是AMP的合成就需要十三个步骤,十二种酶。除了作为基础的ribose-5-phosphpate(5-磷酸核糖)外,还需要5分子的ATP提供能源,一分子的GTP,一分子的二氧化碳,两分子的谷氨胺……
而且,在这十三个步骤中,从第三到第十一步产生的所有中间化合物,除了合成 AMP 或 GMP 以外,都没有别的用处。
在没有套上具体的化合物以前,Horowitz(霍洛维茨)于1945年发表的“A→B→C→D”理论颇具说服力。
他的理论说,最初是直接用D的,后来D快用完了,于是能够利用C→D的物种竞争生存下来;C快用完了时,能够利用B→C→D的生存下来;依次类推,就形成了现在的A→B→C→D。
但这是在没有应用到具体的化合反应以前的空想,当把具体的化合物套进去的时候,一大堆问题出来了。
比方说,几乎每个中间步骤都需要酶的精心控制;一些中间化合物不稳定……等等。这就同样回到了开头那种“需要精心控制反应条件与自然界随机的不可控制条件”之间的矛盾。
五、眼睛瞳孔结构
贝希所举的另一个例证,是人的眼睛在各种外界条件下的自我调整。
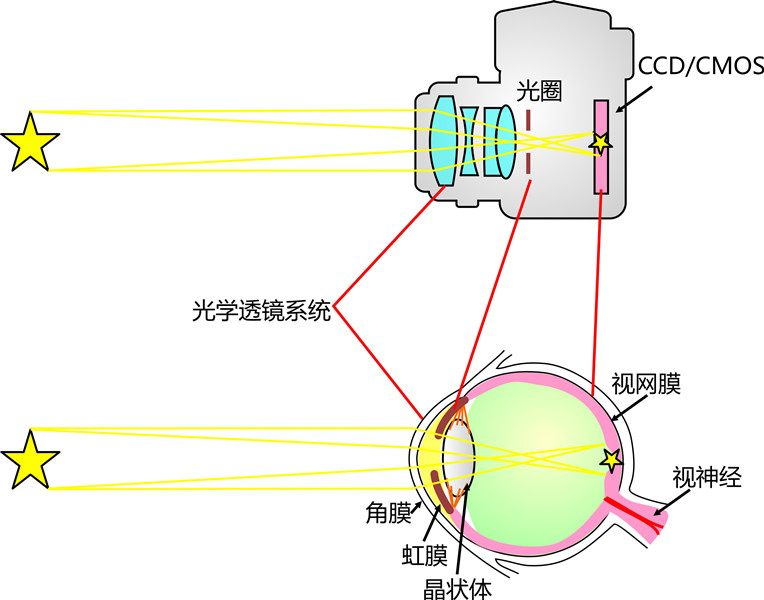
实验证明,人眼的瞳孔有如照相机快门,无论在阳光照耀下或在黑夜里,都能让足量的光线进入眼球,水晶体表面能自动改变光密度,调整色差等等。
这些复杂的机制,连那些自认为熟识眼睛结构的人都惊叹不已。
只是从进化论的角度来解释整个眼睛的解剖结构,是远远不够的。
被达尔文认为如此简单的每一个解剖的步骤和结构,实际上包含着用语言文字难以表达的极为复杂的生化过程。
不论能接受光子作用的细胞与不能接受光子作用的细胞的差别,在达尔文看来是多么“微小”,但在贝希看来却是巨大的——因为接受光子作用的亚细胞结构,是一个具有不可约化复杂性的“分子生化系统”。
达尔文的“自然选择”,显然对此无能为力。
有幸率先看到细胞内部精细结构的贝希教授,用许多无可辩驳的事实告诉世界:
达尔文的进化论是虚浮的,是靠模糊的想象推导出来的。
——进化论即使在常规自然科学的范围,也远不是关于生命起源的最好解释,存在着许多不可逾越的鸿沟。
在旧达尔文主义者看来似乎可以分解成许多步骤的变化,在基因水平上现在必须一蹴而就。而问题是:
你可以跨越一个8英尺宽的沟,但跨越15英尺就让人怀疑。从一个旧物种到一个新物种之间的距离却是100英尺,当达尔文告诉人们,可以分几次跨越过去的时候,人们发现,在这100英尺下面什么也没有!
面对分子水平的毁灭性分析,面对处处断裂,无声反驳的化石链(包括可怕的寒武纪生物大爆炸),许多新达尔文主义者扬弃了达尔文的渐进主张,开始鼓吹激进的“大突变”!
认为不知为什么,不知缘于怎样一种机制,在某一时刻,无数基因会莫名其妙一起突变:只需“嘎嘣”一下,进化过程就完成了,真是神奇到了无以复加,可笑到了极点!
实际上,“大突变”是达尔文曾经深思熟虑,并断然予以拒绝的一种魔术进化论。他说:
“如果我的自然选择论必项借重这种突然进化的过程才能说得通,我将弃之如粪土……如果在任何一个步骤中,需要加上神奇的进步,那自然选择论就不值分文了”。
事实上,进化论的解释,不是由科学的观测事实,而是由“没有上帝”的前提推导而来。
达尔文主义与其说是一种科学,毋宁说是一种哲学。它不外乎是“无神论”的科学包装,利用了基督教人士不知道怎样去证明鬼神存在的致命弱点。
弗朗西斯·培根说得好:“肤浅的哲学使人沦为无神论,而深刻的哲学则把人导向宗教。”
贝希教授所举事例包括了真核生物,意在说明生命自生与进化理论的难以置信。
追本溯源,原核生物的简单,只是相对真核生物而言。从分子层面、化学层面观察,一个最普通的蓝细菌,已经复杂到难以表述。
可惜中国的中学课本对此语焉不详、含糊模棱,一般大学生物学教材内容庞杂,对这部分也是一笔带过、基本忽略。
要真正明确这一点,唯有深入研究生物化学。
翻开清华大学王希成先生编著的《生物化学》,我们可以看到DNA的复制、RNA的合成、蛋白质的合成、核苷酸的代谢、氨基酸的代谢、脂代谢、细胞膜的结构,基因调控等等是何等的繁琐复杂,令人眼花缭乱,心惊神骇。
其中许多生化反应都是原核生物与真核生物所共有的(微生物和多细胞生物,在细胞的化学组成、结构、代谢途径上有着惊人的相似性),完全可以指明某些科学家所谓“生命可以自行合成的假说”是怎样的不近情理。
如果把这些证据一一复制给大家,则最少需要上百页。
而且这也仅仅是细胞工厂中,时时刻刻正在进行着的化学反应的一部分而已。用王先生自己的话说:
“大多数细胞都能进行成百上千个反应,面对这样繁多的反应,如何去描述呢?
本书不可能每一个反应都涉及到,而是主要讨论糖、脂、氨基酸、核苷酸、核酸和蛋白质等生物大分子的代谢反应。”
——仅仅这一部分,王先生就花了四百九十页篇幅!
从物质的分解再加上鞭毛这样一个器官,已经可以使人对最初生命的复杂程度印象深刻。
我们再举一个光合作用的例子吧。
如果一个人无视极多证据,硬要相信生命是自发组织而成、演化而来,我们也没有办法。