前面已经说过,葡萄糖通过糖酵解、柠檬酸循环和氧化磷酸化可以变成二氧化碳和水,同时生成大量的能量货币ATP。对于蓝细菌来说,葡萄糖还需要自己生产。这就要通过“光合作用”,利用捕获到的太阳光量子,将大气中的二氧化碳和水合成为糖。
蓝细菌是目前发现到的最早的光合放氧生物。它启动了纯粹的光合作用,除了叠层岩证据,我们还知道:
一、12C和13C这两种碳同位素是稳定的,其比值在化石的有机物残留中保持恒定,可作为一种光合作用指示剂。
分析表明,在太古代已经具有很好的光合作用了,可能最早的时间是35亿年以前,甚至有可能是在38亿年前。
碳同位素证据与蓝细菌的出现,恰好吻合。
二、释放氧气的光合作用还在极大规模上,永久性地改变了古代岩石的成分和外观。
今天地球上大部分的铁矿石都来自于一组特定的古代岩石中所谓的条带状赤铁矿,形成时间是20 - 35亿年前。
推理可知,那时分子氧首先与铁离子反应,将铁氧化为化合态,并沉积在了海床上,这个过程持续了将近15亿年。用来固定铁的氧气据估计差不多相当于今天大气中氧气的20倍。
当时唯一释放氧气进行光合作用的微生物就是蓝细菌——这也为蓝细菌和光合作用提供了额外的地质学证据(其它的证据就不必提了)。
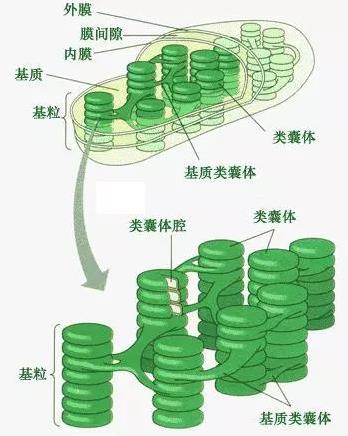
植物和藻类的光合作用发生在自己的细胞器叶绿体中。体内是堆叠的扁平状的颗粒类囊体和其延伸部分—基质类囊体,里面有叶绿素分子和含呈溶解状态的酶。
真核细胞叶绿体:
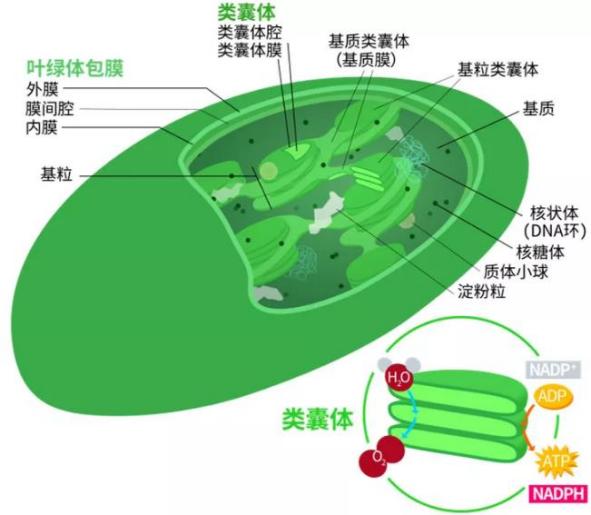
光合作用的第一步是光能被叶绿素吸收并将叶绿素离子化,产生的化学能被暂时储存在三磷酸腺苷(ATP)中,并最终将二氧化碳和水转化为氧气和碳水化合物。叶绿素吸收大部分的红光和蓝光却反射绿光,所以它们会呈现出绿色。
恪守进化论的科学家目前普遍认为,叶绿体是由蓝细菌演化而来,认为它是被原始真核细胞掳入胞内,形成了共生关系。
虽然这是笑谈,亦可见二者结构之相似、之大同小异。
更早的蓝细菌不像真核藻类和植物一样具有叶绿体。其细胞壁内有原生质膜,可分为两个主要区域,其一即是有光合色素的色质区。它主要由类囊体及其有关结构,藻胆体和糖原颗粒等所组成,里面有叶绿素a、藻胆素(藻蓝素、别藻蓝素、藻红素和藻红蓝素)及多种类胡萝卜素分子。
所有色素的共同特征是有亲水的卟啉环,它含有吸收光子的共轭双键网络和疏水的植醇侧链。
蓝细菌具有开创性地以水作为电子来源进行光合作用,固定CO2,放出氧气。与后来的植物一样,进行自养生活。
许多种类的蓝藻细胞壁中甚至含有非光合色素,具有吸收紫外线的功能,可以适应数十亿年前没有保护性气体的地球大气层。
许多蓝细菌的细胞中还有气泡,其作用可能是使菌体漂浮,并使菌体能保持在光线最多的地方。
美国科学家最近还发现,数量很大的海洋浮游蓝细菌,能通过改变酶的种类来充分利用光中的优势波段。
2010年,国外某些研究人员甚至在蓝细菌中发现了一种新的叶绿素分子(被命名为叶绿素f-Chl f),使菌体能够吸收利用近红外光。——最原始的生命啊,其实并不简单。
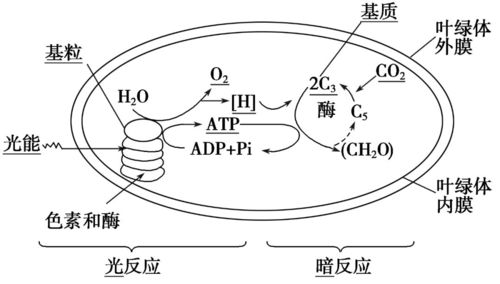
蓝细菌的光合作用部位:
图/蓝细菌光合作用
光反应发生在类囊体膜上,暗反应发生在叶绿体基质中。
蓝细菌与真核生物的光合作用在程序上基本一致,发生的部位不大一样而已。
真核生物的光合色素并不随机分布,而是在类囊体膜中与脂和蛋白质有规律地组成叫做光系统的许多功能单位。一个光系统大约含有250到400个叶绿素和其它色素分子。它有两个主要部分:捕光复合物和光反应中心。光能是通过与每个反应中心相联系的天线色素和称之叶绿素a/b集光复合物的一个色素复合物捕获的。
每个光系统都含有一个反应中心。反应中心由蛋白质复合物(特殊的电子传递分子)和一对特殊的叶绿素a分子组成。光系统中的色素吸收光子,将吸收的光能转移给位于反应中心的特殊的叶绿素分子。只有这个特殊的叶绿素分子直接参与光化学能转换为电化学能的过程,即通过特殊叶绿素吸收的能量可以启动一个电子在电子传递链中的传递,使得光能转换为化学能。
类囊体膜的光系统分两种类型:光系统I(PSI)和光系统II(PSII)。PSI主要位于基质片层,暴露于叶绿体基质;PSII主要位于基粒片层中,远离基质(光系统I比光系统II要原始,但电子传递先在光合系统II开始)。光系统I能被波长700nm的光激发,又称P700。光系统II的吸收高峰为波长680nm处,又称P680。把它们在空间隔开是为了防止这两个光系统之间激发能的自发转移。确保PSI和PSII只通过电子传递联系。
除了光系统,还有一些镶嵌在类囊体膜或者与它相连接的成分也参与光合作用。如细胞色素bf复合物、水裂解复合物、叶绿体ATP合成酶等等。
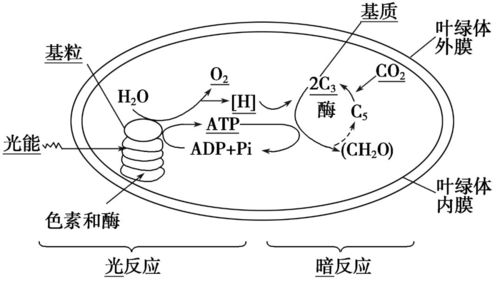
图/光合作用流程
从上图可见:
在光能的驱动下类囊体内进行着电子和质子流动。在水裂解复合物的作用下,来自H2O裂解产生的电子经PSII、一系列中间载体、PSI,最后传递给NADP+;而一部分质子在电子传递过程中从基质泵入类囊体腔内,与H2O裂解生成的质子在腔内外形成一个质子梯度,腔内的质子经ATP合成酶形成的质子通道重新再流回基质,同时催化ATP的合成。
真核植物和蓝细菌的光合作用可分为光反应和暗反应两个步骤:
光反应即电子传递和光合磷酸化。光照强度,水分供给,氧的含量对之皆有影响。两套光合作用系统合作发挥作用。完成了电子传递、水的光解,产氧,将光能转变成化学能,产生ATP,为下一步暗反应提供能量、提供还原剂。利用水光解的产物氢离子,合成NADPH及H离子等等工作。
光照时,天线叶绿素被激发;激发被传递给P680和P700分子。P680和P700分子放出高能电子。电子沿着类囊体膜上的电子传递链传递,经过细胞色素b6/f等等的参与,最后传递给辅酶NADP,通过铁氧还蛋白-NADP还原酶将NADP还原为NADPH。而水光解所得的氢离子则因为顺浓度差通过类囊体膜上的蛋白质复合体从类囊体内向外移动到基质,势能降低,其间的势能用于合成ATP,以供暗反应所用。
电子传递,分循环和非循环两种。
非循环电子传递过程大致如下:
电子从光系统2出发。光系统2→初级接受者(Primary acceptor)→质体醌(Pq)→细胞色素复合体(Cytochrome Complex)→质体蓝蛋白(含铜蛋白质,Pc)→光系统1→初级接受者→铁氧化还原蛋白(Fd)→NADP+还原酶(NADP+ reductase)
非循环电子传递链会裂解水,释出氧气,生产ATP与NADPH。
循环电子传递过程概略如下:
电子从光系统1出发。光系统1→初级接受者(Primary acceptor)→铁氧化还原蛋白(Fd)→细胞色素复合体(Cytochrome Complex)→质体蓝蛋白(含铜蛋白质)(Pc)→光系统1
循环电子传递不会产氧,因为电子来源并非裂解水。最后也会生产出一些ATP。
光合作用中消耗的ATP比NADPH要多得多,因此当ATP不足时,相对来说会造成NADPH的累积,刺激循环式电子流之进行。
再看一遍,类囊体膜上的非循环电子传递链:
光合作用的实质是把CO2和H2O转变为有机物和把光能转变成ATP中活跃的化学能再转变成有机物中的稳定的化学能。即光能→电能→不稳定的化学能(能量储存在ATP的高能磷酸键)→稳定的化学能(合成为淀粉等糖类)。
这就需要下一步的碳反应。利用光反应形成的ATP和NADPH,将CO2还原为糖。
“碳反应”原来错误的被称为“暗反应”,在20世纪90年代的一次光合作用会议上,科学家们已经一致同意,将暗反应改称碳反应。
碳反应可分为多种类型,最常见的是C3类型。
“C3循环”又称“卡尔文循环”,是光合作用里暗反应的一部分。反应场所为真核生物叶绿体内的基质。
循环可分为三个阶段:
羧化、还原和二磷酸核酮糖的再生。大部分植物,会将吸收到的一分子二氧化碳,通过一种叫“二磷酸核酮糖羧化酶”的作用,整合到一个五碳糖分子1,5-二磷酸核酮糖(RuBP)的第二位碳原子上。此过程称为二氧化碳的固定。这一步反应的意义是,把原本并不活泼的二氧化碳分子活化,使之随后能被还原。但这种六碳化合物极不稳定,会立刻分解为两分子的三碳化合物3-磷酸甘油酸。后者被在光反应中生成的NADPH+H还原,此过程需要消耗ATP。产物是3-磷酸丙糖。
后来经过一系列复杂的生化反应,一个碳原子,将会被用于合成葡萄糖而离开循环。剩下的五个碳原子经一系列变化,最后再生成一分子1,5-二磷酸核酮糖,循环重新开始。
循环运行六次,生成一分子的葡萄糖。
具体过程如图:
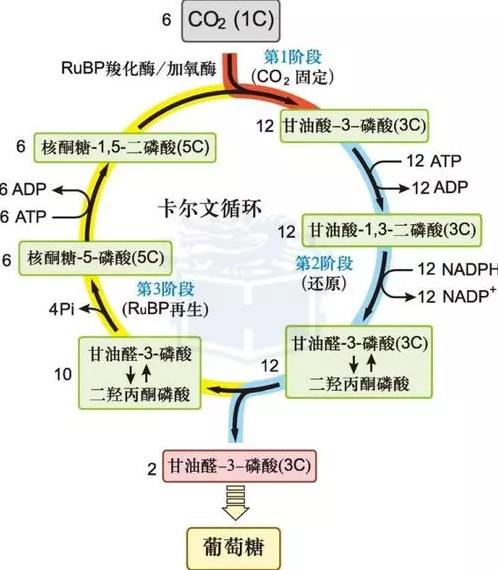
图/卡尔文循环
所有使用卡尔文循环进行碳固定的细胞,在进行光合作用的同时,还要进行光呼吸,包括蓝细菌。这是在有光照及细胞中高氧低二氧化碳的情况下发生的,之所以称为光呼吸(又叫氧化的光合碳循环;乙醇酸途径或C2旁路),是因为这一过程会吸收氧气放出二氧化碳。
光呼吸会抵消约30%甚至更多的光合作用,对生物似乎大为不利。
光呼吸与光合作用伴随发生的根本原因被认为主要是由细胞中世界上最丰富的酶—Rubisco—1,5 - 二磷酸核酮糖羧化/加氧酶的作用决定的。因为它既可催化羧化反应,又可以催化加氧反应,也就是说CO2和O2要竞争Rubisco酶的同一个活性部位,两种情形既相互抑制又相互促进。
在光呼吸过程中,参与卡尔文循环的反应物1,5-二磷酸核酮糖和催化剂1,5-二磷酸核酮糖羧化酶/加氧酶发生了与其在光合作用中不同的反应。光合作用中,二磷酸核酮糖在羧化/加氧酶的催化下增加两个氧原子,再经过一系列反应,最终生成3-磷酸甘油酸。后者再经过部分卡尔文循环中的步骤,可再次重新生成为二磷酸核酮糖。但在光呼吸过程中,二磷酸核酮糖在羧化/加氧酶的催化下生成2 - 磷酸乙醇酸。
光呼吸和卡尔文循环密不可分。
人们把它们之间的关系形象的理解为:糖工厂内(行卡尔文循环的细胞)的葡萄糖生产线(卡尔文循环)因一部机器(1,5 - 二磷酸核酮糖羧化酶/加氧酶)构造不完善,一部分原材料(1,5 - 二磷酸核酮糖)不断被错误加工,产出次品(2 - 磷酸乙醇酸),虽然有一补救措施,可将次品重新加工并再次投入生产线,但是整个过程却是非常费时费力的。这个错误加工和补救的过程就是光呼吸。
发生光呼吸的细胞需要三个细胞器的协同作用才能将光呼吸起始阶段产生的“次品”“修复”,耗时耗能。早先光呼吸被人们称作“卡尔文循环中的漏逸”,“羧化/加氧酶的构造有缺陷”。后来科学家才发现,光呼吸可消除多余的还原型辅酶II(NADPH)和ATP,回收碳素,维持光合碳循环。防止强光下过量生成的超氧阴离子自由基对光合系统、光合色素的伤害,保护细胞。因为光呼吸与大气中氧气/二氧化碳比例联系非常紧密,科学家甚至认为可以通过控制陆地植物的数量,以控制地球大气氧气和二氧化碳的成分比。
在真核细胞中,光呼吸发生的场所为叶绿体,过氧化物酶体和线粒体。有人认为,像蓝藻这种水生的原核生物,有卡尔文循环发生的结构和其地点羧化体,但是却没有过氧化物酶体和线粒体,光呼吸即使会发生,似乎也只能进行到乙醇酸这一步。蓝藻的细胞膜上有碳酸根泵,可以从周围介质中主动吸收无机碳并积累之。提高羧化体中的二氧化碳浓度。而高浓度的二氧化碳会压制光呼吸。
所以,在20世纪80年代有人怀疑,究竟蓝藻中是否会发生光呼吸。——目前大抵认为蓝细菌肯定有光呼吸,但又能有效压制光呼吸。
目前这个问题似乎还难以搞清楚。光呼吸和光合作用是在大气中存在光照条件下同时进行的,加上细胞本身会进行呼吸作用,一般的气体交换方法难以发现和测定光呼吸。人们对于各种中间产物在各细胞器中的转运和光呼吸的调节,还所知甚少。蓝细菌压制光呼吸,与玉米、高粱和甘蔗等C4植物一样高效率进行光合作用也许是一件好事。可以看到:目前的许多研究者正在准备让C3植物向蓝细菌等“低等生物”取经学习。
光呼吸过程非常复杂,在真核生物中,涉及三个细胞器的相互协作:叶绿体、过氧化物酶体和线粒体。
光呼吸的开始部分,叶绿体部分:
1分子氧气能与1分子1,5-二磷酸核酮糖生成1分子2-磷酸乙醇酸和3-磷酸甘油酸。反应由1,5-二磷酸核酮糖羧化酶/加氧酶羧化/加氧酶催化。这1分子磷酸乙醇酸会被磷酸乙醇酸磷酸酶脱去磷酸机团成为乙醇酸。乙醇酸在叶绿体内膜上有相应的转运体,它协助乙醇酸离开叶绿体。乙醇酸到达过氧化物酶体时,会通过可能是由孔蛋白组成的孔进入过氧化物酶体。
而横向来看,光呼吸的最终阶段也是发生在叶绿体。由过氧化物酶体得来的甘油酸会转变为3-磷酸甘油酸,而后者也是光呼吸开始时二磷酸核酮糖分解和卡尔文循环羧化阶段的产物。3-磷酸甘油酸会进入卡尔文循环余下的两个阶段:还原阶段(产物是丙糖磷酸)和1,5-二磷酸核酮糖再生阶段。
同时,叶绿体也能将α-酮戊二酸还原为谷氨酸。这是光呼吸过程中谷氨酸-酮戊二酸循环中的一部分。再生的谷氨酸会再回到过氧化物酶体内与乙醛酸进行转氨基作用。
过氧化物酶体部分:
过氧化物酶体的基质是细胞中处理有毒物质的特殊场所。但通过对拟南芥的研究,过氧化物酶体具有比以前认为的(即脂类降解,光呼吸和过氧化氢解毒三大作用)更多功能。在光呼吸过程中产生的乙醛酸和过氧化氢(双氧水)都是有毒害作用的物质。即使该两种物质低浓度的存在于叶绿体中,也能够完全阻断光合作用的发生。原因是,乙醛酸和过氧化氢会氧化卡尔文循环中硫氧还蛋白的二硫键,硫氧还蛋白因此失去激活下游蛋白的能力。乙醛酸还能抑制羧化/加氧酶。
在过氧化物酶体中,乙醇酸加氧成为乙醛酸,并生成过氧化氢。
过氧化氢会被过氧化物酶体中的过氧化氢酶催化为水和氧气。而乙醛酸也会在谷氨酸的参与下通过转氨基作用生成甘氨酸,催化的酶是谷氨酸乙醛酸转氨酶。甘氨酸通过孔道逸出过氧化物酶体到达线粒体,通过转运进入后者参加下一步反应。
而在线粒体生成的丝氨酸则又会回到过氧化物酶体,这时的丝氨酸会作为氨基供体,通过丝氨酸乙醛酸氨基转移酶转变为羟化丙酮酸,后者在NADH供氢的情况下被还原为甘油酸,返回叶绿体。而丝氨酸乙醛酸氨基转移酶和谷氨酸乙醛酸转氨酶所催化的反应都是植物体内调节氨基酸含量的重要过程。
与线粒体和叶绿体膜的选择性通透不同,过氧化氢和乙醛酸非常容易通过过氧化物酶体膜逸出。但这并未发生,是因为过氧化物酶体的基质的特殊性质。实验发现,倘若线粒体或叶绿体的膜被破坏(例如将两者悬浮于水中,所谓的“渗透休克”即会发生细胞膜破裂),线粒体和叶绿体的内容物会溶解。但过氧化物酶体的内容物在膜破裂后却会以颗粒状存在,颗粒大小与原过氧化物酶体相当。这说明,在过氧化物酶体中,酶是以复合体的形式结合在一起的。一系列的酶促反应在复合体中各个部分之间能快速传递,又能防止底物逸出和副反应的发生,是一种非常高效的代谢形式,被称为“代谢物沟道效应”。
线粒体部分:
在线粒体中,两分子的甘氨酸会在甘氨酸脱羧酶复合体的作用下脱去一分子二氧化碳和氨,生成一分子丝氨酸。
这一步反应其实是非常复杂的。甘氨酸脱羧酶复合体由含硫辛酰胺辅基的H蛋白,含磷酸吡哆醛辅基的P蛋白,含四氢叶酸的T蛋白和L蛋白组成。参与反应的一分子甘氨酸首先与P蛋白的吡哆醛上的醛基反应,生成一分子施夫碱。甘氨酰残基然后会被脱羧(除去-COO?),只剩下-CH2NH3+,再后会被带到H蛋白的硫辛酰胺残基上,这是一步氧化还原反应,其中硫辛酰胺的二硫键被还原。之后T蛋白参与反应,断开碳原子和氮原子之间的连接。氮元素以氨的形式释放。而碳原子则被T蛋白转移到另一甘氨酸的α碳原子上,成为一分子丝氨酸。
反应中生成的NADH能够被线粒体呼吸链用作能量的生成,同时也能作为还原当量被供给其他细胞器利用。绿色植物线粒体具有很强的甘氨酸氧化能力,其甘氨酸脱羧酶复合体可占线粒体中溶解蛋白质的30到50%。非绿色植物的甘氨酸氧化蛋白含量则很少,甚至缺失。
——我们摘录细胞中的一小部分化学反应,只是为了给平常不涉足生物化学的网友们留下一个印象:
细胞代谢的全过程的确是十分复杂的。
一个小小的细胞,哪怕肉眼看不见,却是一座效率极高的微型化工厂。
许多人误以为,“复杂的生物化学反应,是从真核生物—真核细胞才开始的”。
许多这方面的书刊,直截了当地胡说,“最早的原核生物十分简单”,致令许多不求甚解的人产生了“生命可能来自物质自组织”的荒谬思想。
通过上面有限的几个例子,透过冰山一角,我们已经看到最原始的生命极其复杂。这还仅仅是从人的角度来查考。
在神的眼里,原核细胞还有更复杂、更奇异的结构(碍于禁令我们只好避而不谈),它的下面还有基本粒子中的更小世界更小世界,一层又一层,就像俄罗斯套娃(已经不在我们的探讨范围)。
伟大的尼古拉·特斯拉(Nikola Tesla)即曾指出:
粒子的生命比人类生命要更加复杂上百万倍。